CHAPTER 04
효소
20개 시트1,364개 노드표 2개그림 179개교재 2종 통합전문가 정밀 구조
Obsidian 통합 · 전문가 재구성
효소반응 속도론
효소1
효소반응 속도론5
- Michaelis-Mentene 식
가수분해8
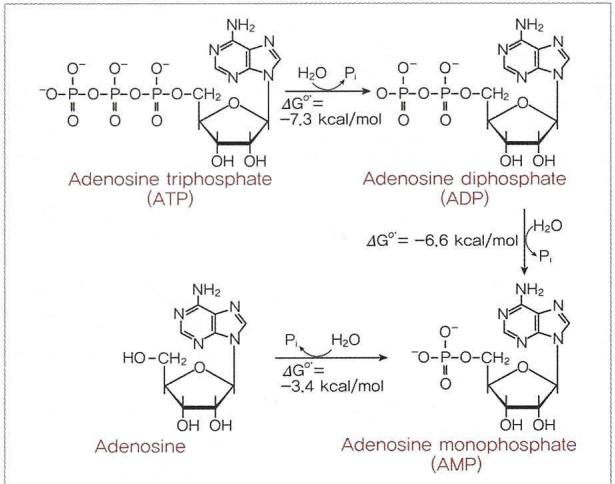
- ATP는한 분자의 무기인산(HOPO32,p)을 방출하여 ADP가된다이과정을발열반응이며표준조건하에서가수분해도ㄴ1M의 ATP 당 7.3kca의 에너지가방출된다
- 세포내의조건은 표준조건과 다르므로 세포는 7.3kcal보다 많은 양의 에너지를 방출한다.
- ATP의인산은음으로하전되고서로반발하므로불안정하다.
- 따라서인접한 인산과의 공유결합을 유지하기위해서는많은에너지를 포함하고있을것이며 인산기의 가수분해시많은 에너지를 방출하게 된다.
- 활성화된 세포는초당 수백만 개의ATP를요구한다.
- 평균적으로한분자의AP는만들어진 후1\2분이내에소비되고다시 형성되므로 휴식 중인 인간은하루 약 40\60kg의 ATP를생성 및 소비하며 평균 체내 약 57\100g 정도가 존재한다.
- 전형적인세포는ADP 한 분자 당 1O개 이상의ATP 분자들을가지고있으므로AP 가수분해 반응이훨씬 더잘이루어진다.
 가수분해
가수분해
Lineweaver-Burk 식2
- V = K m V m a x \cdot 1 [ S ] + 1 V m a x
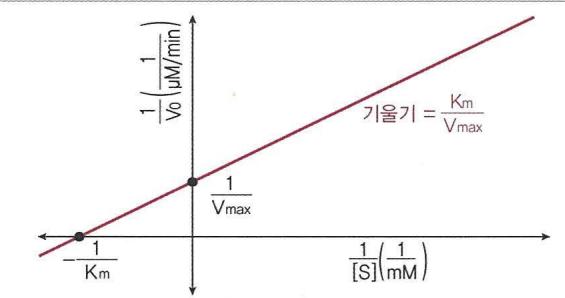 Lineweaver-Burk 식
Lineweaver-Burk 식
- 기능
효소반응과 관련된 여러 상수41
- K m (미카엘리스 상수)
- 정의 1 2 V m a x 지점에서의기질 \eqcirc
- 초기속도 V 0 ) > t 최대속도( V m a x ) 의절반이되는지점에서 K m \underline o [S]와 같다.
- V m a x 2 = V m a x [ S ] K m + [ S ] 1 2 = [ S ] K m + [ S ]
- \therefore K m = [ S ] > 1 된다.
- ㄴ 특징
- K m ^ \circ 활성자리의절반이채워져 \therefore \bigcirc \sqrt \equiv 때의 기질의농도이다.
- 따라서 K m 은의미있는촉매작용이 일어나는데 필요한 기질 농도의 척도가 된다.
- b K m 값은 효소에 따라서 매우 다르며, 또한 같은 효소라도기질pH,온도 등에 따라서다르다.
- k - 1 \gg \ k 2 \stackrel \circ = 경우에한해서 K m 은ES 복합체의 친화력 즉, 효소와 기질의 친화력(affinitly)을 나타낸다.
- 만약 k - 1 \gg k 2 \equiv 1 \sqcup
- K m = k - 1 + k 2 k 1 \ \longrightarrow \ K m = k - 1 k 1 이며
- 정상 상태에서k\[ES] = k 1
- E][S]이ㅁ로
- K m = k - 1 k 1 = [ E ] [ S ] [ E S ] = k d
- 즉. k - 1 \gg k 2 일경우 K \mathfrak m \mathcal \supseteq [ES]의 해리상수와 같다.
- 따라서큰 K m 값은[ES]복합체의 약한 결합을 의미하며 작
- 은 K m 값은[ES]복합체의 강한 결합을 가리킨다.
- 즉. K m 은효소와 기질의 친화도를 의미하게 된다.
[S] < K m 경우기질 농도 증가와 반응속도는비례적으로증가한다.1
- 1차 반응양상
[S] > K m 경우기질 농도 증가와관계없이 반응속도는 일정하다.1
- 0차 반응 양상
k c a t1
- 대사회전수, 교체수(turnover number)
- k c a t \bigcirc \bigstar 효소1분자의활성(mdlecular activity)이라고도하며효소가기질로포화되었을때 효소1분자에의해단위시간당생성물로 변화하는기질수를의미한다.
- 보통의효소반응의경우MM식의가정처럼두단계로만구성되지않으므로일반적인속도상수인 k c a t 을정의한다.
- 만약반응이및단계로구성되며 그중하나가속도제한단계인경우 k c a t 은그속도제한단계의 속도상수와 같다.
- \therefore k c a t = k 2
- V m a x = k 2 [ E t ] = k c a t [Et]이다.
- : k 2 = k c a t = V m a x [ E t ]
k e a t / K m1
- 촉매효(catalytic efficiency)
- 생리적조건하에서대부분의 효소는기질에포화되어 있지 않다. 즉 [S]< K m 이며대부분의활성지리는점유되어있지 않고 효소반응의 속도상수는 k c a t ( k 2 ) 보다훨씬작다.
- 따라서 속도상수 k c a t ( k 2 ) 와효소농도로는 실제 반응을예측하기어렵기때문에 k c a t / K m 0
- 제안되었다
- 매우낮은기질농도를지닐경우의 반응속도식
- ) \odot \downarrow [ \ V = k 2 [ E S ] = k c a t [ E S ] \ \ [ E S ] = V k c a t
- 식 [ES] = [E] [ S ] k m 을[ES]에 대해 정리
[ E S ] = V k c a t = [ E ] [ S ] K m1
- \ : V = k c a t K m [ E ] [ S ]
- 매우낮은 기질 농도에서는대부분의 효소가 효소기질복합체보다는 자유효소로 존재하므로
- \therefore V = k c a t K m [ E t ] [ S ]
- k c a t / K m \geq 속도상수로 K m ^ \circ 효소의기질친화력에반비례하고 k c a t 은효소의 속도에정비례하므로 k c a t / K m 은기질농도가 포화농도보다 훨씬 못 미치는조건에서 작용하는 효소의 촉매효을(efficiency
- )에 대한지표를 제공한다.
- 따라서 매우 낮은 기질 농도에서 반응속도는효소농도[Et], 기질 농도[S] 그리고 k e a t / K m 에직접적으로비례하며반응은V와S가직선관계이로1차반응이다.
효소반응 속도론
효소
효소반응 속도론
Michaelis-Mentene 식
가수분해
ATP는한 분자의 무기인산(HOPO32,p)을 방출하여 ADP가된다이과정을발열반응이며표준조건하에서가수분해도ㄴ1M의 ATP 당 7.3kca의 에너지가방출된다
세포내의조건은 표준조건과 다르므로 세포는 7.3kcal보다 많은 양의 에너지를 방출한다.
ATP의인산은음으로하전되고서로반발하므로불안정하다.
따라서인접한 인산과의 공유결합을 유지하기위해서는많은에너지를 포함하고있을것이며 인산기의 가수분해시많은 에너지를 방출하게 된다.
활성화된 세포는초당 수백만 개의ATP를요구한다.
평균적으로한분자의AP는만들어진 후1\2분이내에소비되고다시 형성되므로 휴식 중인 인간은하루 약 40\60kg의 ATP를생성 및 소비하며 평균 체내 약 57\100g 정도가 존재한다.
전형적인세포는ADP 한 분자 당 1O개 이상의ATP 분자들을가지고있으므로AP 가수분해 반응이훨씬 더잘이루어진다.
Lineweaver-Burk 식
V = K m V m a x \cdot 1 [ S ] + 1 V m a x
기능
효소반응과 관련된 여러 상수
K m (미카엘리스 상수)
정의 1 2 V m a x 지점에서의기질 \eqcirc
초기속도 V 0 ) > t 최대속도( V m a x ) 의절반이되는지점에서 K m \underline o [S]와 같다.
V m a x 2 = V m a x [ S ] K m + [ S ] 1 2 = [ S ] K m + [ S ]
\therefore K m = [ S ] > 1 된다.
ㄴ 특징
K m ^ \circ 활성자리의절반이채워져 \therefore \bigcirc \sqrt \equiv 때의 기질의농도이다.
따라서 K m 은의미있는촉매작용이 일어나는데 필요한 기질 농도의 척도가 된다.
b K m 값은 효소에 따라서 매우 다르며, 또한 같은 효소라도기질pH,온도 등에 따라서다르다.
k - 1 \gg \ k 2 \stackrel \circ = 경우에한해서 K m 은ES 복합체의 친화력 즉, 효소와 기질의 친화력(affinitly)을 나타낸다.
만약 k - 1 \gg k 2 \equiv 1 \sqcup
K m = k - 1 + k 2 k 1 \ \longrightarrow \ K m = k - 1 k 1 이며
정상 상태에서k\[ES] = k 1
E][S]이ㅁ로
K m = k - 1 k 1 = [ E ] [ S ] [ E S ] = k d
즉. k - 1 \gg k 2 일경우 K \mathfrak m \mathcal \supseteq [ES]의 해리상수와 같다.
따라서큰 K m 값은[ES]복합체의 약한 결합을 의미하며 작
은 K m 값은[ES]복합체의 강한 결합을 가리킨다.
즉. K m 은효소와 기질의 친화도를 의미하게 된다.
[S] < K m 경우기질 농도 증가와 반응속도는비례적으로증가한다.
1차 반응양상
[S] > K m 경우기질 농도 증가와관계없이 반응속도는 일정하다.
0차 반응 양상
k c a t
대사회전수, 교체수(turnover number)
k c a t \bigcirc \bigstar 효소1분자의활성(mdlecular activity)이라고도하며효소가기질로포화되었을때 효소1분자에의해단위시간당생성물로 변화하는기질수를의미한다.
보통의효소반응의경우MM식의가정처럼두단계로만구성되지않으므로일반적인속도상수인 k c a t 을정의한다.
만약반응이및단계로구성되며 그중하나가속도제한단계인경우 k c a t 은그속도제한단계의 속도상수와 같다.
\therefore k c a t = k 2
V m a x = k 2 [ E t ] = k c a t [Et]이다.
: k 2 = k c a t = V m a x [ E t ]
k e a t / K m
촉매효(catalytic efficiency)
생리적조건하에서대부분의 효소는기질에포화되어 있지 않다. 즉 [S]< K m 이며대부분의활성지리는점유되어있지 않고 효소반응의 속도상수는 k c a t ( k 2 ) 보다훨씬작다.
따라서 속도상수 k c a t ( k 2 ) 와효소농도로는 실제 반응을예측하기어렵기때문에 k c a t / K m 0
제안되었다
매우낮은기질농도를지닐경우의 반응속도식
) \odot \downarrow [ \ V = k 2 [ E S ] = k c a t [ E S ] \ \ [ E S ] = V k c a t
식 [ES] = [E] [ S ] k m 을[ES]에 대해 정리
[ E S ] = V k c a t = [ E ] [ S ] K m
\ : V = k c a t K m [ E ] [ S ]
매우낮은 기질 농도에서는대부분의 효소가 효소기질복합체보다는 자유효소로 존재하므로
\therefore V = k c a t K m [ E t ] [ S ]
k c a t / K m \geq 속도상수로 K m ^ \circ 효소의기질친화력에반비례하고 k c a t 은효소의 속도에정비례하므로 k c a t / K m 은기질농도가 포화농도보다 훨씬 못 미치는조건에서 작용하는 효소의 촉매효을(efficiency
)에 대한지표를 제공한다.
따라서 매우 낮은 기질 농도에서 반응속도는효소농도[Et], 기질 농도[S] 그리고 k e a t / K m 에직접적으로비례하며반응은V와S가직선관계이로1차반응이다.